The number and variety fishes that may be found in the stream or river near your house may surprise you. Most, of course, are not game fish. If you live in the United States, the kind of fish you find will depend largely on what region you are from. Fish communities change from one area to another because of such factors as water temperature, water velocity and clarity, alkalinity, and available habitat. For instance, a typical Pacific mountain stream may contain the fry and smolts of salmonids (trout and salmon), sculpins (fishes of the Family Cottidae), various small minnows that like shallow and rapidly-moving water, and brook lamprey. A typical Great Basin stream, on the other hand, being muddier, deeper, and warmer, may contain catfish, suckers, many minnows, and members of the Family Centrarchidae, with bass and sunfish of various kinds.
Amazingly, however, rivers all over the world have fishes that are closely related. From the North America to Europe to Russia to China, you can still find salmon, eels, sturgeon, minnows, and suckers that would seem quite familiar to you.
Streams in the United States are home to many large fishes that are so well known to us that they won't be discussed here: pike, gars, eel, sturgeons, bowfin, lamprey, salmon, trout, catfish, bass, sunfish, and shad are just a few. There are other fishes, though, that are not as well known because they are harder to see and find.
Most of the fishes in a typical U.S. stream are in the family Cyprinidae--that is, they are minnows. The common Cyprinids are carps, shiners, daces, minnows, and chubs. Cyprinids are omnivorous. They move along eating mostly from the bottom (bottom-feeders), and will select anything from algae to insects to worms. Some minnows only do well in slow or stagnant waters. Others have a wonderful hydrodyamic shape. Their bodies and fins are shaped in a way that causes the water flowing over them to push them downward onto the riverbed. Cyprinids like these prefer riffles and rapids--shallow, fast-flowing water. Their mouths tend to be pointed downward (subterminal), for more efficient feeding off the bottom. The dace and the chub, shown below, are examples of these adaptations.

|

|
| long-nosed dace | speckled chub |

|
| shiner |

|

|
| darter | logperch |

|
| sculpin |

|
| white sucker |

|
| killifish |
In Pacific mountain streams, which are rocky and clear, fishes tend to live on the bottom, in "holes" or deep pools below boulders and woody debris, or under ledges or debris. The salmonid fry that have been excluded by competition and are forced to live in the open nevertheless sport a color-scheme that ensures they will blend in with the bottom.
This is not the only way fishes adapt to different regions. For example, fishes in a clear stream tend to be like salmonids: visual predators, and capable of swimming large distances at high speed, but not very maneuverable. Fishes in a muddy stream tend to be those that can either find their food by filtering the water or shoveling or sucking from the bottom, or those that are highly maneuverable in close quarters, like centrarchids. There are exceptions, of course. For every stream there are fishes that have evolved to take advantage of every available habitat niche.
Since fishes have evolved to inhabit almost every kind of watery habitat on Earth: those which dry up during drought; mountain streams and deep, muddy rivers; stagnant pools and vast lakes; freezing polar regions and tropical seas; the unknown, darkest depths of the ocean; underground caves and shallow tidepools; salt water, fresh water, and often both--you might expect that their biological diversity, the variations in their bodies and behaviors, would be mind-boggling. You would be correct!
Fish biologists, in an attempt to categorize the many fishes of the world so that they can communicate about them to other biologists, most often use morphology (the study of physical types) to separate the different types of fishes. One can separate fish by body type, air bladder type, fin type, scale type, and mouth type, to list just a few possibilities. Other differences used to classify fishes fall more into functional categories, like reproductive type and mode of movement. These distinctions are then used by taxonomists to assign fishes to scientific categories like Class, Family, Genus, and Species.
If you would like to know more about types, click on any of the buttons below:
An interesting thing about these kinds of classifications is that they can also help us figure out how long the fish has been around Planet Earth. For example, if a fish's pectoral fins are fairly low down on the body and stick out horizontally much like a plane's wings (like a shark, salmon, or catfish), we might guess that fish to be more "primitive" than one who sports pectoral fins high up on its body and turned in a vertical direction (like a parrotfish, bass, or sunfish). We would call the second fish more "derived," by which we suggest that it is a later development in the evolution of fishes.
Another interesting benefit of classifications is that they can help us figure out the ecological role that the particular fish we are studying plays in its environment. For instance, one can often tell from a fish's morphology what it eats, how it maneuvers, how and where it hides, how it reproduces, and how much oxygen it needs, and many other things about the fish. The "primitive" fish in the last paragraph we might guess is more of a traveller, and perhaps even a predator on smaller fishes. The "derived" fish, by contrast, might well be a resident of a complex habitat like rock piles or a coral reef, and spend much of its time darting in and out of hiding places. Why? Because low, horizontal fins are of use for little more than guidance and balance, while high, vertical fins can be used by the fish to swim backwards, stop suddenly, and turn "on a dime."
The relationships in aquatic communities are often complex due to the presence of so many different organisms filling the many different habitat niches available in such bodies of water. Nevertheless, morphological characteristics of the fish present can tell us a great deal about the intricately woven fabric of each community. Biologists can tell a great deal about the attributes and needs of each community merely by looking at the morphology of the fishes present, as well as types of plants and animals and physical characteristics of the stream.
The relationships in aquatic communities are usually poorly understood only because we lack the money and other resources to thoroughly study each aquatic community on Earth. Unfortunately, we humans seem to have very little interest in knowing about such things. You can see for yourself what can happen when humans go about using aquatic resources to make money without understanding the ecology that is involved. Click on the button below to see what happened in Jamaica.
The devastation of Pacific salmon on the West Coast of North America is well worth studying. These fishes went from being a primary resource for native americans to being on Threatened and Endangered status (Endangered Species Act of 1973) within a little over one century of European influence. What happened? Few have described it better than Jim Lichatowich in his 1999 book, Salmon Without Rivers. I recommend it.
Follow the Ecology link below for more information on fishes, or go directly to the Trout and Salmon page.
 Fish Biology (brief but thorough)Saving Endangered Fishes
Fish Biology (brief but thorough)Saving Endangered Fishes 
| Site Search |
There are many different body types, and several different ways of describing them. What follows is a very simple summary of a few body types. Below the pictures you will find their names and descriptions.
| Sagittiform | Taeniform |
|---|---|

|
|
| "Arrow-like." Example shown is a grass pickerel. Other fish with this body type include pikes, gars, topminnows, killifish, needlefish, and barracuda. These body shapes are good for rover predators, which depend on an ability to strike quickly--often from a hiding place. | "Ribbon-like." Example shown is a gunnel. This shape is good for hiding in cracks and crevices, but fishes like this do not move very fast. |
| Depressiform | Fusiform |

|

|
| "Flattened." Flounders, halibut, rays, and skates all have this form, which allows the fish to rest on the bottom and hide either using camouflage or by covering itself with a layer of sand. In streams, sculpins come close to having this shape. | Streamlined and shaped like a plane fuselage. The example shown is a blue-fin tuna. Tunas, mackerel, swordfish, sailfish, and marlin are all fusiform fishes. This shape allows them to move through the water extremely fast. In streams, most minnows have fusiform shapes. |
| Compressiform | Anguilliform |

|
|
| Compressed laterally. Example shown is a green sunfish. This shape is highly versatile and is probably the most common fish shape. It combines the advantages of several of the other fish body shapes. Many freshwater fishes have this shape, such as bass, crappie, and sunfish. | "Eel-like." Many eels, of course have this shape. Shown is a brook lamprey. This shape allows a fish to enter and hide in very narrow openings, and also helps the fish resist the force of current. |
| Filiform | Globiform |

|

|
| "Filament-like." Shown is a snipe eel. This would be an unusual shape in a freshwater environment. | "Globe-like." Shown is a smooth lumpsucker. This, too, would be an unusual shape in a freshwater environment, although pupfish come close. |
Another way to classify body types is by their function. In this system:
1. Rover predators (fish that more or less constantly swim about searching for prey) include fusiform body types, as well as salmon, trout, and bass; they have pointed heads, terminal mouths, narrowed caudal peduncles, and forked tails.
2. Lie-in-wait predators (predators that catch their prey by ambush) include sagittiform body types; they have dorsal and anal fins placed well back on the body, a streamlined form, flattened heads, and large, well-toothed mouths.
3. Surface-oriented fishes are often small, with mouths that are directed upwards toward the surface of the water. These fishes often swim just below the surface, and eat food that is floating on the surface or flying above it. Topminnows, killifish, freshwater hatchetfish, halfbeaks, and flying fish are examples of surface-oriented fishes.
4. Deep-bodied fish include all the compressiform types, and are found widely in places where the ability to make tight, close turns is of value, such as rock reefs, coral reefs, thickly vegetated areas, and schools.
5. Eel-like fish include taeniform, anguilliform, and filiform shapes all in one category.
6. Bottom fish is a very broad category that includes bottom rovers like catfishes, suckers, and sturgeons, bottom clingers like sculpins, bottom hiders like darters and blennies, and depressiform body types, like flatfish.
You can see that this functional method is very descriptive and helpful. Still, it leaves out a number of fish. What about a puffer? A pipefish? A seahorse?
Most fishes (but not all) have air bladders, which we will call gas bladders here. The primary function of the gas bladder is to allow the fish to achieve neutral buoyancy; that is, to keep from sinking. To achieve neutral buoyancy--that is, to be able to stay at any depth it wants--a fish has to be able to take gas into the bladder and let gas out of it, just like you might blow up a balloon or let it deflate.
There are two major different types of gas bladder in fishes: physostomous, in "primitive," and physoclistous, in "derived" fishes.
Fish with physostomous air bladders take gas into their bladders using their mouths: they rise to the surface and swallow air. To let out air, they just burp it out. Examples of this kind of fish are catfish, trout and salmon, sturgeon, minnows, suckers, and eels.
Physoclistous gas bladders, however, do not open to the mouth, so the fish has to let gas in and out of the bladder using a very complex little patch of blood vessels that absorb or let go of gases from the blood. Fishes with these bladders include bass, perch, mosquitofish, and sunfish.
Since physoclistous fishes cannot just burp the gas out of their bladders, they cannot change depths quickly. This is why many fishes, when taken quickly from deep habitats to the surface, will eventually die: they cannot deflate their bladders quickly enough. A good example of this is the rockfish. Many rockfish are caught by nets but fisherman do not want them because of young age, small size, or because they were not looking for rockfish: these fish, even when released, will likely die. (This is unfortunate, because rockfish are long-lived fish, often living to 80 years or more. Because they live a long time, they do not reproduce in large numbers like other fish. Therefore, once you have killed the small population of rockfish in one area, it may be gone forever).
There are other ways for fishes to store gases, and fishes use their gas bladders for other things besides buoyancy, such as respiration, hearing (all minnows), and sound production. Deepsea fishes often store fat in their gas bladders rather than gases. One, the Coelacanth, stores fat in lungs--for the purpose of bouyancy.
Before we start talking about fins, let us be clear about where they are. The diagram of the strange, non-existent fish below will help you follow the discussion
of fin types.
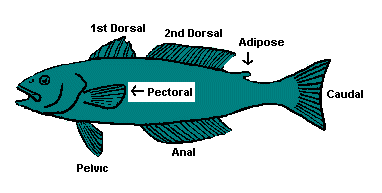
Spines:
Spines are a late development in fishes, and many fish have them. Spines may be found almost anywhere on the body, and are used primarily for defense. Spines may have painful poisons in them, such as those of the catfish, or simply be annoying because they are sharp and serrated. Some fishes have developed large spines that merely make it difficult for other fishes to get their mouths around them, or impale the unfortunate diner. The most common locations for spines are the first part of the dorsal fin (or in fishes with two dorsal fins, the first dorsal fin), and the first part of the pectoral fins. Spines may be counted and their location noted, and this often helps us tell one species from another. Spines may also be cut and their rings counted to provide the fish's age, much like a forester would age a tree trunk.
Pectoral fins:
Pectoral fins may be horizontal and down low, like in a salmon, trout, shark, or sturgeon, and used mainly for gliding. Over the course of millennia they have moved upwards on the body of many fishes. Most reef and schooling fishes have their pectoral fins up high and oriented vertically, as in the above diagram. These are often are used for swimming, holding position, and changing directions quickly. Some fishes have further specialized their pectoral fins; the sculpin uses its broad, spiny pectoral fins to help it hold itself to the bottom against strong currents. Flying fishes have lengthened their pectoral fins to allow them to glide. Some fishes use only their pectoral fins to swim, sculling them through the water like oars.
Pelvic fin location:
Pelvic fins are usual abdominal, meaning that they are attached midway down the belly. Salmon and trout have pelvic fins at this location. When the pelvic fins are below the pectoral fins, such as can be seen in the diagram of the non-existent fish above, they are termed thoracic. When a thoracic pelvic fin is attached under the gills, it may also be called jugular, and if under the chin or eye, mental.
Caudal fin shape:
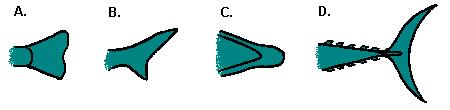
A. The homocercal (homo-SUR-kul) tail is a modern development. It is symmetrical. It includes truncate, square, slightly forked, and deeply forked types. It is by far the most common caudal fin shape, shared by most fishes.
B. The heterocercal tail is an ancient form, possessed by only a few primitive fishes, such as sharks, sturgeon, and paddlefish. It was a necessary tail shape when fishes had no swim bladders and were heavy in the front; if the fish tried to use a symmetrical tail, it would have plunged toward the bottom. Instead, it developed a tail with a deliberately downward-driving design and supplemented it with horizontal, plane-like pectoral fins that transformed that downward force into a horizontal, forward-driving force. Genius!
C. This tail has a non-differentiated caudal fin. This may be found on eels of all sorts, as well as lampreys.
D. The lunate tail is a tail that the tuna and mackerel have refined to an engineer's delight. This tail provides powerful forward force, with very little turbulence.
As you might expect, this short list only grazes the very surface of the subject of fin shapes and types, and how they may be used to distinguish one fish from another! Not only fin shapes and types may be used, but also the number and length of rays, the number and length of spines, and the spatial relationship of one fin to another.
Scales have evolved over time and are of major importance in classifying fishes. Most scales are deeply buried in the fish's epidermis, or outer skin layer, with only part of them showing. Below the pictures of scales are examples of how the scales would look on the fish's body.
There are four common types of scales in fishes. They are shown in the figure below. Below each scale type is an illustration of what they would look like on the fish.
| Ganoid | Placoid | Cycloid | Ctenoid |
|---|---|---|---|

|

|

|

|
| This "primitive" kind of scale is an evolutionary reminder of the time when fishes used armor plating to protect themselves. Ganoid scales are hard and smooth, and may take the form of only a few scales (or scutes, as in the sturgeon and stickleback), partial plating, or overall body plating. Sturgeons, gars, and sticklebacks have ganoid scales (or scutes). | Sharks have placoid (PLAK-oyd) scales: tiny, tooth-like structures that are partially embedded in the skin. These tiny, pointed scales, made of the same materials as their (and our) teeth, make their skin feel like sandpaper. | Many fishes with which we are most familiar have cycloid scales, which are the thin, round, almost transparent scales that we find when we are cleaning trout, salmon, or herring. Minnows also have cycloid scales. These scales are mostly buried in the epidermis, allowing only the small posterior margin to show. | Highly evolved fishes often have ctenoid (TEEN-oyd) scales, which are much like cycloid scales except that they have tiny, comb-like projections (ctenii) on their posterior edges (the edges that show, and are not buried in skin). The colors of brightly colored fishes also show on these posterior edges. |
Besides the above types, there are also cosmoid scales, as well as scaleless fishes (sculpins, many catfish, some eels, and swordfish), and fishes which have scales so deeply buried that they look scaleless (many tunas and anguillid eels).
Scales appear to have originated as bony outgrowths from the skin. The gar's ganoid scales still have layers of bone, along with enamel and ganoine. The shark's placoid scales, however, have become nearly identical to our own teeth, with a layer of enamel over dentine and a pulp cavity. Cycloid and ctenoid scales have evolved in another direction entirely. Scientists have long been puzzled by the shininess of fish scales. Recent x-ray analysis has demonstrated that these scales are built from stacks of collagen fibrils laid down parallel in layers, just like plywood. Each layer of fibrils is oriented at a 36 degree angle from the one beneath it, giving five directions in all. Tiny hexagonal apatite crystallites orient themselves parallel to the fibers. The combination of the plywood structure and the aligned crystals causes the silvery effect of scales by refracting light multiple times. This silvery shininess, however, is only part of the equation. We also tend to think of a fish's iridescence as "shiny," too. Many fishes have layers of iridocytes beneath their scales and even in their skin. These cells contain tiny granules that act as mirrors. Reacting with pigment cells, iridocytes can give a fish a rainbow effect of different colors that sets off its bright silver as it swims into the light.
You might have noticed that fish that are sick are not shiny like healthy fish. This is because fishes need to maintain their scales and iridocytes by depositing guanine in the iridocytes and by making new scales, and they cannot do so when they are sick. Dead fish are dull, too, becoming duller the longer they have been dead, because the collagen and guanine are proteins that break down after death.
Because salmonids have been introduced to streams around the world, from Costa Rica to New Zealand, most people are probably familiar with the most common salmonid reproductive cycle: adults migrate upstream once a year, sometimes from the ocean; the female digs gravel nests (redds) and lays eggs while the male fertilizes them; the female covers them back up and guards them for awhile, then dies; the hatched fry float back downstream.
As wondrous as this method of reproduction is, however, it is only one of the many, many forms employed by fishes. To begin with, although salmonids are usually much like humans, with nearly a 1:1 ratio of males to females, this is not always so in the fish world. In many reef fishes, there are only a few males to be found. And some go through sex changes, depending on age and necessity. A fish may spend her entire life as a female, only to change to a male in order to "take over" the harem, changing colors, size, and behavior to fit her new role (sequential hermaphroditism) This is common with wrasses, groupers, and parrotfishes. Some fishes are a combination of both sexes at once (synchronous hermaphroditism).
Some fishes have only females; they reproduce without sperm (parthenogenesis), or in one case--a Mexican molly--mate with another species. In this instance, however, the male's genetic material is not used; the offspring are all females, and true clones of the mother. The male serves only to stimulate parthenogenesis. This successful species throws into doubt the oft-repeated maxim that two parents are necessary for the genetic health of a species.
Also, whereas one can usually tell a male salmonid from a female during spawning, many fishes are impossible to sex without sacrificing and dissecting them. Generally, male fishes only stand out from the crowd when they have territories to defend, like a reef or a redd. Obviously, a male herring in a vast school of herring in the ocean has nothing to defend, and thus no reason to be any different from a female.
Deepsea anglerfishes have a bizarre form of reproduction: the male, once he meets a female, becomes attached to her belly. Over time, he gradually loses more and more of himself, and his body's tissues unite more and more with his host--the female--until he is little more than a sack of sperm attached to his mate. Thus, apparently once they have found each other, they do not need to look for mates ever again.
There are different ways of fertilizing as well: some fishes spawn into nests or cavities, some into mouths, some into the open ocean, and some practice internal fertilization. Eggs may be fertilized outside the body, fertilized inside the body and then extruded, or fertilized inside the body and hatched inside the body, as happens in humans. These latter types of fishes are called live-bearers, because they give birth to living fishes.
Finally, there are different ways of caring for the young. As noted above, female salmonids often remain and guard their eggs until they die. Some fishes never give another thought to their eggs. Others take the eggs into their mouths and care for them there even through hatching. Many females guard nests, but so do some males; the male stickleback lures females to a nest that he builds, and then guards the nest until they hatch. A cousin of his, the male seahorse, takes the female's eggs into his own pouch and becomes pregnant with the young.
Some minnows migrate upstream, like salmon, to spawn. They do not spawn in nests, but in the open water. Their eggs float downstream, hatch, and continue to float downstream as fry, until they are large enough to take up residence in a part of the stream. The Rio Grande silvery minnow is one of these. There used to be other fishes that spawned in this fashion in the Rio Grande, but they have all become extinct due to the presence of so many dams and diversions. Obviously, with a fish of this kind, if you make it impossible for them to swim upstream and float downstream, they will eventually go extinct. The situation of the Rio Grande silvery minnow has become complicated by the fact that in modern times a growing population of people is living in the desert. They dry up the Rio Grande every summer in order to water fields of alfalfa (a water-hungry crop), wash cars, and water lawns. Although the fish need upstream access to spawn, their need for any water at all is rather obvious.
Many fishes are identified by looking at or into the mouth. The number of mouth-types exhibited by different species is nothing short of astonishing. Three lakes in Africa contained about 900 species of cichlids, nearly all differentiated mainly by the way their mouths are shaped. (This number is rapidly dwindling, by the way, as the cichlids in these lakes are driven to extinction). Cichlid mouths in Lakes Victoria, Malawi, and Tanganyika are variously adapted to eat other cichlids' eggs, scales pulled from fishes' living bodies, algae from rocks, tiny invertebrates, and many other forms of food. The arrowana of South America has a mouth adapted for spitting water with precision at insects perched on overhead branches. Parrotfish mouths have evolved to look and act like beaks, which they use to grind at coral, making the sand that surrounds coral reefs. Seahorses and pipefish have tubular mouths for sucking in small prey in narrow places like a vacuum cleaner.
Besides mouths, fish also use part of their gills for eating: the gill rakers protrude into the mouth cavity. They can be toothy, small and nubby, or long and comb-like. Long, comb-like gill rakers are useful for filter-feeders, which feed by swimming along with their mouths open, filtering small organisms from the water. Paddlefish, anchovies, and menhaden are good examples of filter-feeders. Toothy gill rakers, on the other hand, can help hold onto unwilling prey.
Fish may have no teeth or different kinds of teeth, as well as teeth in different places in their mouths. Fishes may have jaw teeth, or no jaw teeth. A cutthroat trout may be differentiated from a rainbow trout by the presence of basibranchial teeth, which are just behind the base of the tongue. Loaches and minnows, which have no jaw teeth and look like they are pouting, have plenty of pharyngeal, or throat, teeth for grinding up their food. Fishes may also have teeth on the roofs of their mouths (vomer and palatine teeth).
The positioning of the mouth is important. Most fish-eating fishes have terminal (at the absolute front of the fish's head) mouths. Suckers and other bottom-feeders have subterminal mouths (below the absolute front tip of the head). Surface-feeders have upturned, or supraterminal, mouths.
Possibly the most prominent difference that can be seen is between gape-and-suck feeders and hit-and-run feeders, both of which can be seen in predatory fish. Hit-and-run feeding was an ancient development for fishes. These fishes, like salmon, trout, gar, and barracuda, have large, hinged jaws, most often with some quite sharp teeth. They lunge at their prey, snap their mouths closed around it, and hold onto it with those sharp teeth. This is a classic sort of predation, and works quite well, but modern fishes have improved upon it. Gape-and-suck feeders have developed protrusible mouths; that is, their mouths open much, much wider, and much, much faster, than you could ever have thought possible from the size of their mouths at rest. This is due to a special arrangement of the bones that make up their mouths. These fishes, like bass, sunfish, and rockfish, actually create a kind of vacuum when they open their mouths, sucking water--along with prey--right into their mouths. The advantage of this is that the fish does not have to move quiet so fast or snap so accurately to catch its prey. Some fishes, like scorpionfishes and anglerfishes, do not have to move at all. They merely lurk in their hiding place until a likely fish drifts by, then open their giant mouths and suck them in.
Fishes may move in ways that we don't usually think they can, like certain fishes that can fly, glide, hop, or walk using their pectoral fins. And of course, some fishes dig down into the substrate, or bottom. But even when they are swimming, fishes exhibit a remarkable variety of ways of moving.
Most fishes swim by pushing their bodies against the water sideways--first to one side, and then to the other. The reason for this is the way their bodies' muscles are built, limiting them to lateral movements.
| Anguilliform | Subcarangiform | Carangiform | Thunniform | Ostraciiform |
|---|---|---|---|---|
| (ang-WIL-i-form) | (SUB-ca-RANJ-i-form) | (ca-RANJ-i-form) | (TOO-ni-form) | (oh-STRAY-kee-i-form) |
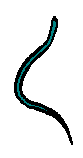
|
|
|

|

|
| The way most eels and many other fishes, such as brotula, gunnels, oarfish, and nurse sharks move. In addition, the sea snakes of Australia's seas swim this way. In streams, eels and lampreys use anguilliform movement. The fish moves the entire body against the water in a snake-like fashion. This method of swimming is surprisingly efficient. Electric eels do not swim this way, but instead hold their bodies very still and undulate only their fins to swim. This is because of the electrical fields that they generate. | This movement category includes salmon, trout, catfish, minnows, and cod. These fishes undulate most of their bodies, but leave their heads fairly still and concentrate most of the movement in the last two-thirds of their bodies. | These fishes include drums, jacks, and snappers, and are very fast swimmers. In streams, bass, crappie and sunfish use carangiform movement. The forward parts of their bodies are not capable of undulation. They throw their bodies into a shallow wave that increases in amplitude as it moves backward toward the tail, which snaps like a whip. As you can see, this leaves the head almost still. The caudal peduncles (the part of the tail just before the caudal fin) are narrow and the tails are often forked, leading to more efficient movement. | This term is sometimes used to describe the extremely fast and efficient movements of fishes like tuna, billfish, and lamnid sharks. These fishes have very narrow caudal peduncles and large caudal fins that are lunate (look like the sliver of a waxing moon). Their tails are reinforced by keels, for strength and stability. This movement type can also be seen in whales and dolphins (mammals), and was used by the extinct ichthyosaurs (reptiles). | Many fishes are hampered by a boxy or globular shape. Trunkfishes, cowfishes, and boxfishes have too much armoring to be able to flex their bodies. These fishes scull their tails like oars. |
Coral must be cleaned constantly of algae, or it will suffocate and die. Algae grows quickly in the tropical sun, and many coral reef fish serve their reef in this way, tirelessly removing every bit of algae from the coral so that it can receive sunlight, clean water, and nutrients. Thus does coral continue to thrive.
Jamaica's coral reefs, like all coral reefs, host fabulously complex communities of organisms. Jamaicans have always fished for and eaten fish from these reefs. But recently, Jamaica's population has swelled. There are many more people who must eat and many more who have no jobs. Because of this, the island's reefs are used as a resource. (Being a tropical island, after all, it has few other resources).
Unfortunately, coral reef fish in Jamaica have been fished so aggressively in order to both provide food for residents and aquarium specimens for aquarists in wealthier nations that there are not enough of them left to graze algae off of the coral. As a consequence, Jamaican reefs are slowly being killed beneath a blanket of algae: more than 90% of its reefs are gone. The algae makes the habitat unusable to other coral species, which then desert the reefs. Is this the end of the story? That is doubtful, because each coral reef's influence spreads far beyond its actual location.
Jamaica's coral reef story is, of course, much more complicated than what I have just described. Factors adding to Jamaican coral destruction have been climate change, sea urchin disease, recreational damage, and sewage from the over-populated island. A good question to ask is, "Would the fish have been able to control the increased algae, had they been left on their reef to do their job?"
This kind of tragedy is not isolated to Jamaica, nor to coral reefs, but is being repeated again and again all over our planet as humans continue to extract resources before knowing enough about the positions of those resources in Earth's intricate webs of life.
 Go back to Inhabitants
Go back to Inhabitants